El retículoendoplasmático es el orgánulo más grande de muchas células eucariotas. Está formado por túbulos (tubos) y sacos (cisternas) que se comunican formando una red continua desde la membrana nuclear, extendiéndose por todo el citoplasma de la célula. La parte interna del retículo endoplasmático se llama lumen (o espacio luminar o cisternal).
La membrana del RE tiene estructura similar a la plasmática, aunque es algo más delgada y tiene menos lípidos y más proteínas que la membrana plasmática.
Según sus funciones y su composición se distinguen dos tipos de retículo endoplasmático:
Retículo endoplasmático rugoso (RER)
Tiene ribosomas adheridos en sus membranas, en el lado del citosol. Los ribosomas están unidos por su subunidad mayor por la ayuda de unas proteínas del grupo de las riboforinas, que no se encuentran en el REL.
Se comunica con la membrana nuclear y el retículo endoplasmático liso.
Está formado por sacos y grandes cisternas aplanadas.
Su función principal es la de síntesis de proteínas con los ribosomas fijados a su membrana. Las proteínas sintetizadas pasan del RER al REL, luego al aparato de Golgi y de allí, a los lisosomas, a la membrana plasmática o al exterior. En el recorrido por estos orgánulos, la molécula experimentará un procesado adicional.
Las proteínas sintetizadas y almacenadas en el RER, antes de ser transportadas a otros orgánulos citoplasmáticos (aparato de Golgi, lisosomas), a la membrana plasmática o al exterior de la célula, deben ser glucosiladas para convertirse en glucoproteínas.La glucosilación de las proteínas se produce en el lumen del retículo, mientras que las proteínas del citosol no suelen estar glucosiladas.
Retículo endoplasmático liso (REL)
Carece de ribosomas, por lo que su superficie es lisa.
Formado, mayoritariamente, por una red de túbulos, que se unen al RER, que se extiende por todo el citoplasma.
La mayor parte de las células tienen un retículo endoplasmático liso escaso, pero es especialmente abundante en:
Células musculares estriadas.
Células intersticiales ováricas de Leydig, del testículo y células de la corteza suprarrenal que segregan hormonas esteroideas.
Hepatocitos, donde participa en la síntesis de moléculas lipoproteicas.
Funciones del retículo endoplasmático liso:
Síntesis, almacenamiento y transporte de lípidos.
En el lado citoplasmático de su membrana se sintetizan casi todos los lípidos constituyentes de las membranas: colesterol, fosfolípidos, glucolípidos, etc.
Los ácidos grasos se sintetizan en el citosol y entran en la membrana por acción de una flipasa.
En algunas células, el retículo endoplasmático liso produce grandes cantidades de lípidos, como por ejemplo, las hormonas esteroideas.
Contracción muscular. En las células del tejido muscular estriado, el REL libera calcio activando la contracción muscular.
Detoxificación. Su membrana contiene enzimas desintoxicantes que degradan sustancias liposolubles que puedan resultar nocivas y las transforman en sustancias solubles que pueden ser excretadas por el organismo. En los vertebrados, la detoxificación se realiza principalmente en las células del hígado, riñones, intestino, pulmones y piel. Por ejemplo, el REL abunda en las células del hígado, donde contiene enzimas que desintoxican productos nocivos como el alcohol y otros subproductos del metabolismo, como el amoníaco.
Liberación de glucosa a partir del glucógeno (en hepatocitos). El glucógeno almacenado en el hígado se encuentra en forma de pequeños gránulos adheridos a las membranas del REL. Cuando se necesita energía, el glucógeno se degrada obteniéndose glucosa-6-fosfato en el citoplasma. El REL elimina el grupo fosfato y las moléculas de glucosa entran en el REL y, son enviadas a la sangre hacia donde se requieran.
Produce vesículas de transporte con proteínas y lípidos recién sintetizados para llevarlos hacia el aparato de Golgi.
Aparato de Golgi
Aparato de Golgi con sus sectores
El aparato de Golgi es un orgánulo presente en todas las células eucariotas, excepto en los glóbulos rojos. Pertenece al sistema de endomembranas. Está formado por unas estructuras llamadas dictiosomas (dependiendo del tipo de célula), y estos dictiosomas están compuestos por unas 4 - 8cisternas (sáculos discoidales) aplanadas rodeadas de membrana que se encuentran apiladas unos encima de otras, y cuya función es completar la fabricación de algunas proteínas. Funciona como una planta empaquetadora, modificando vesículas del retículo endoplasmático rugoso. El material nuevo de las membranas se forma en varias cisternas del aparato de Golgi
Las células eucarióticas poseen un aparato de Golgi más o menos desarrollado, según la función que desempeñen. Los dictiosomas pueden presentar continuidad con otros componentes del sistema de endomembranas, como por ejemplo el retículo endoplasmático.
Estructura y composición del aparato de Golgi
Los dictiosomas presentan:
Una cara CIS, proximal o de formación: es la más interna, cercana al núcleo, y continuación de las membranas del retículo endoplasmático, del que recibe las vesículas de transición, que son sáculos con proteínas que han sido sintetizadas en la membrana del retículo endoplasmático rugoso (RER), introducidas dentro de sus cavidades y transportadas por el lumen hasta la parte más externa del retículo. El contenido de las vesículas de transición se incorpora a las cisternas del aparato de Golgi. De cada cisterna salen nuevas vesículas que se incorporan a la siguiente cisterna.
Su forma es convexa. Sus membranas son finas, de composición similar a la de las membranas del retículo endoplasmático.
Una cara intermedia o sáculos de la zona central, que tienen la mayor actividad metabólica. Presenta las vesículas intermedias o intercisternas.
Una cara TRANS, distal o de maduración: se encuentra próxima a la membrana plasmática (se parece más a ella). Su forma es cóncava. En la periferia de esta cara, se encuentran unas vesículas grandes llamadas vesículas de secreción, que contienen en su interior los productos finales y que llevarán a la membrana plasmática para expulsarlos al medio externo (exocitosis) o para formar lisosomas, que contienen enzimas digestivas.
Funciones del aparato de Golgi
Transporte y concentración de proteínas (formadas principalmente en el RE). Las proteínas procedentes del RER, englobadas en vesículas se unen a la región cis del dictiosoma. Aquí, si no están fosforiladas, las proteínas sufren una fosforilación. Las proteínas secretadas se van desplazando, pasando de una cisterna a otra, hasta llegar a las cisternas situadas en la cara trans del dictiosoma.
Glucosilación de lípidos y proteínas. Se completa la glicosilación que había comenzado en el RE. En el aparato de Golgi se produce el ensamblaje de oligosacáridos a lípidos y proteínas para formar glucolípidos y glucoproteínas respectivamente. También se fabrican los glucosaminoglucanos de la matriz extracelular de las células animales, así como las pectinas y la hemicelulosa de las paredes de las células vegetales.
Formación del del fragmoplasto en células vegetales. El tabique telofásico que divide el citoplasma en la zona ecuatorial de la célula vegetal, se produce por la asociación de vesículas derivadas del aparato de Golgi.
Formación del acrosoma en el espermatozoide. El acrosoma deriva del aparato de Golgi. Contiene enzimas hidrolíticas que sirven para digerir los componentes de las cubiertas del óvulo durante la fecundación.
Formación de membranas (cuando se expulsa al exterior el contenido de las vesículas mediante exocitosis, se unen a la membrana plasmática, incrementando la superficie).
Formación de lisosomas.
Concentración de sustancias (glúcidos, proteínas). Recolector y transporte.
Cuando se han sintetizado las glucoproteínas y los glucolípidos, irán a la membrana plasmática a través de las membranas de secreción.
Las proteínas (formadas en el RER) y los lípidos (formados en el REL) son transportados al aparato de Golgi por las vesículas de transición y allí van pasando de sáculo en sáculo hasta salir por las vesículas de secreción transformados en glucolípidos y glucoproteínas.
Los dictiosomas son estructuras muy dinámicas, ya que los sáculos del aparato de Golgi se renuevan constantemente por fusión de las vesículas de transición procedentes de las membranas del RE, y se van transformando los sáculos hasta llegar a ser vesículas de secreción.
Lisosomas
Los lisosomas son orgánulos globulares, formados por el aparato de Golgi, presentes tanto en células animales como vegetales.
Los lisosomas son vesículas membranosas, en cuyo interior tiene lugar la digestión intracelular. Su membrana es de tipo unitaria de 75 Å, recubierta por el interior por una gruesa capa de glucoproteínas que la protegen de la destrucción por la acción de las enzimas hidrolasas. La membrana del lisosoma contiene una bomba de protones que introduce H%2b y mantiene el pH ácido, inferior a 5, en el interior del lisosoma, y proteínas de transporte que permiten que pasen los productos resultantes de la digestión al citosol.
El lisosoma es el marcado con el número 12
Composición química y tipos de lisosomas
Los lisosomas contienen unas 50 enzimas diferentes del tipo hidrolasas ácidas, sintetizadas en el retículo endoplasmático rugoso. Son enzimas capaces de digerir las macromoléculas y tiene su funcionamiento óptimo en un medio con un pH = 5. La membrana del lisosoma contiene proteínas ácidas integrales altamente glucosiladas, que la protegen de las enzimas que contiene.
Los lisosomas contienen una gran cantidad de enzimas hidrolíticas, pero su contenido puede ser muy distinto. Se distinguen dos tipos de lisosomas:
Lisosomas primarios (también llamados “vesículas con hidrolasas”). Sólo contienen enzimas hidrolíticos. Son vesículas de secreción recién formadas por gemación a partir de las cisternas del aparato de Golgi. Pueden formar lisosomas secundarios o expulsar su contenido al exterior.
Lisosomas secundarios. También contienen hidrolasas, pero además sustratos en vía de digestión. Son lisosomas primarios fusionados con otras sustancias, de origen interno o externo. Según la sustancia que hidrolizan, se distinguen dos tipos:
Vacuolas heterofágicas: fusión de un lisosoma primario con partículas que proceden del exterior de la célula (fagolisosoma).
Vacuolas autofágicas: fusión de un lisosoma primario con distintas partes de la célula que ya no son necesarias o zonas lesionadas (autofagolisosoma).
Funciones de los lisosomas
Los lisosomas son los encargados de la digestión de las macromoléculas, aunque en algunos casos, sirven como almacén temporal de reservas:
1.- La digestión celular. Los lisosomas pueden digerir macromoléculas, tanto de origen interno como externo. Esta función la realizan de dos formas:
Autofagia. La materia que digiere es una parte de la célula, como fragmentos del RE, sáculos o cisternas del aparato de Golgi. La autofagia es importante para las células, ya que destruye zonas dañadas, interviene en el desarrollo celular y permite la nutrición de la célula cuando las condiciones son desfavorables.
Heterofagia. Lo digerido tiene origen externo. Tiene la doble finalidad de nutrir y defender la célula. Los sustratos entran a la célula por endocitosis, formando una vesícula que se fusiona a un lisosoma primario, originando a una vacuola digestiva, donde se produce la digestión. Los productos de la digestión pasan al hialoplasma, donde son reutilizados. Lo que no puede ser digerido, se expulsa al exterior por exocitosis, pudiendo fusionarse con la membrana plasmática.
2.- Almacenamiento de sustancias de reserva. En las semillas existe un tipo especial de lisosomas secundarios, los granos de aleurona, que almacenan las sustancias de reserva. No se produce la digestión celular hasta que no llega el momento de la germinación. Entonces se activan las enzimas hidrolíticas, iniciando la digestión intracelular, cuyos productos se utilizarán en el desarrollo del embrión.
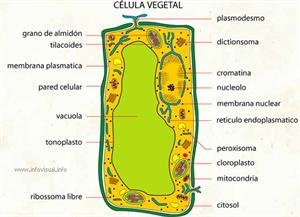
Vacuolas
Vacuolas en rosa
Las vacuolas y las inclusiones son lugares del interior de la célula en los que se acumulan sustancias. Si están rodeados por una membrana se denominan vacuolas; en caso contrario, inclusiones.
Son orgánulos rodeados por una membrana simple, con forma redondeada y que contienen sustancias que no están involucradas en ningún proceso metabólico. Se forman en células jóvenes, por fusión de vesículas derivadas del retículo endoplasmático y del aparato de Golgi.
Las células eucariotas animales tienen bastantes vacuolas. Las células eucariotas vegetales, cuando son jóvenes tienen muchas vacuolas, pero cuando son maduras pueden llegar a tener una única célula que ocupa el 90% del volumen celular.
Dentro de la vacuola se encuentra el jugo vacuolar amorfo, formado por agua y que puede contener diversas sustancias de almacén o de desecho.
El conjunto de vacuolas de una célula recibe el nombre de vacuoma.
Tipos de vacuolas
Vacuola vegetal
Las células vegetales jóvenes tienen muchas vacuolas pequeñas porque contienen poca agua, pero según van creciendo, aumentan de tamaño y se fusionan formando una gran vacuola central que ocupa casi todo el citoplasma.
Las vacuolas vegetales son generalmente incoloras (excepto las de los pétalos de las flores). La membrana que forma vacuola vegetal se llama tonoplasto.
La principal función de las vacuolas en vegetales consiste en almacenar sustancias.
Vacuola contráctil (vacuola pulsátil)
En las células animales, las vacuolas están relacionadas con el aparato de Golgi, con funciones de almacenamiento de sustancias, transporte, etc. Ya hemos hablado de ellas cuando vimos los lisosomas: vacuolas digestivas y vacuolas autofágicas.
Entre las vacuolas animales destacan las vacuolas pulsátiles que presentan las células que viven en ambientes hipotónicos, como es el caso de muchos protozoos, y que son utilizadas para regular la flotabilidad bombeando el exceso de agua al exterior por transporte activo.
Funciones de las vacuolas
Las funciones principales de las vacuolas son:
Mantenimiento de la turgencia celular. La presión osmótica dentro de las vacuolas es muy alta por la elevada concentración de sustancias. El agua tiende a penetrar en las vacuolas por ósmosis, lo que provoca que la célula se mantenga turgente, contribuyendo a que la planta esté erguida.
Permiten que aumente de tamaño la célula vegetal, sin que ello suponga un excesivo gasto de energía por su parte.
Almacenamiento de sustancias diversas. Las vacuolas pueden servir como almacén para muchas moléculas de la célula:
Agua.
Sustancias de reserva, como glúcidos, ácidos grasos y proteínas.
Productos de desecho.
Pigmentos, como antocianinas y flavonas de los pétalos de las flores.
Sustancias de desecho resultantes del metabolismo celular y que pueden resultar tóxicas para la planta si están libres en el citoplasma. Por ejemplo, la nicotina o el cianuro.
Perixosomas
Estructura básica del perixosoma
Los peroxisomas son unos orgánulos esféricos, pequeños, presentes en células animales y vegetales, rodeados de una membrana unitaria de 75Å, que contienen unas enzimas oxidasas, entre las que destacan la peroxidasa y la catalasa.
Los peroxisomas son unos orgánulos muy parecidos a los lisosomas, pero se diferencian de ellos en que no contienen hidrolasas ácidas, sino enzimas oxidativos.
Son abundantes en células que sintetizan, almacenan o descomponen lípidos.
En los peroxisomas se realizan reacciones de oxidación similares a las producidas en las mitocondrias, pero la energía producida se disipa en forma de calor, en lugar de almacenarse en forma de ATP.
La peroxidasa utiliza oxígeno molecular para oxidar diversos tipos de sustratos desprendiendo peróxido de hidrógeno (H2O2), sustancia que es tóxica para la célula. Después actúa la catalasa, que descompone el peróxido de hidrógeno.
Así, pueden oxidar ácidos grasos y aminoácidos, aportando energía para la célula, además de detoxificar una gran variedad de moléculas tóxicas, sobre todo en el hígado y en el riñón.
Las funciones de los peroxisomas son:
Detoxificación de algunas sustancias tóxicas para el organismo, como ácido úrico, etanol, metanol, …
Degradación de ácidos grasos y aminoácidos que no van a generar ATP, como ocurre en la mitocondria, sino energía en forma de calor.
Gluxisomas
Un tipo especial de peroxisomas son los llamados glioxisomas, exclusivos de células vegetales, que aparecen en semillas en germinación y permiten producir glúcidos a partir de los ácidos grasos.
Mitocondias
Las mitocondrias están presentes en casi todas las células eucarióticas, mientras que los cloroplastos sólo aparecen en las células vegetales fotosintéticas. Los dos se incluyen dentro de los orgánulos energéticosde las células.
Si no fuera por las mitocondrias, las células heterótrofas sólo podrían obtener su ATP de la glucólisis anaerobia, pero con ellas pueden degradar totalmente la glucosa en presencia de oxígeno hasta convertirla en dióxido de carbono y agua, obteniendo mucha más energía.
El número de mitocondrias depende de las necesidades energéticas de la célula, situándose en zonas en las que se requiere un consumo importante de ATP.
Estructura de la mitocondrias
Las mitocondrias tienen forma cilíndrica y están limitadas por una doble membrana, la membrana mitocondrial externa, que la separa del hialoplasma, y la membrana mitocondrial interna, que tienen unos repliegues hacia el interior que aumentan su superficie, las crestas mitocondriales. Estas membranas dejan dos compartimentos: el espacio intermembrana, entre ambas membranas, y la matriz mitocondrial, espacio que rodea la membrana mitocondrial interna.
Funciones de las mitocondrias
La función principal de las mitocondrias es la obtención de energía, aunque cada componente mitocondrial tiene distinta función:
El hecho de que los compartimentos mitocondriales tengan distintos complejos enzimáticos, implica que en cada uno de ellos se realizarán funciones diferentes:
Ciclo de Krebs. Tiene lugar en la matriz mitocondrial.
Cadena respiratoria. En el ciclo de Krebs se desprenden una serie de electrones. Los transportadores de electrones se localizan en la membrana mitocondrial interna.
Fosforilación oxidativa. Se realiza en las crestas mitocondriales. La ATP-asa fosforila el ADP y lo transforma en ATP. Así se sintetiza la mayor parte del ATP que se produce en las células aerobias.
La β-oxidación de los ácidos grasos. Las enzimas que intervienen en ella están situadas en la matriz mitocondrial.
Concentración de sustancias en la cámara interna, tales como proteínas, lípidos, colorantes, hierro, plata, calcio, fosfatos y partículas semejantes a los virus.
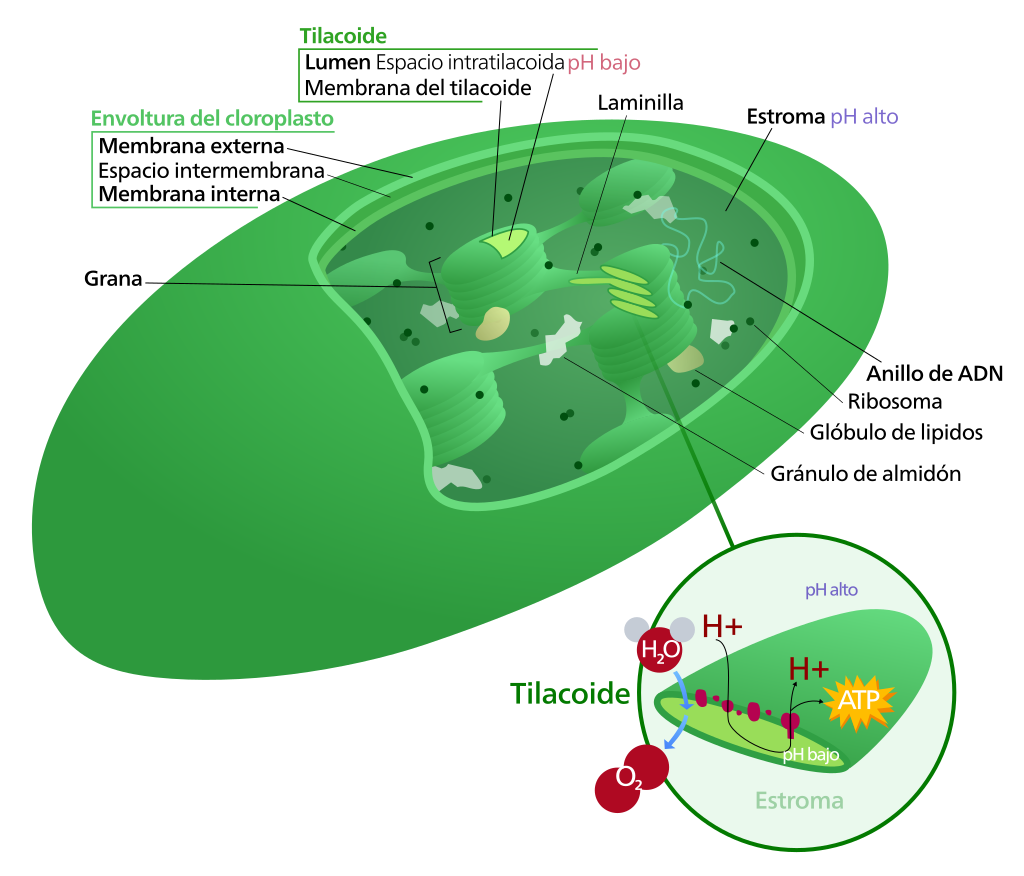
Cloroplasto
Los cloroplastos son un tipo de cromoplastos que se localizan en las células vegetales fotosintéticas y en las algas verdes.
Se pueden mover por movimientos ameboideos y contráctiles, y se colocan en la parte de la célula que recibe mayor cantidad de luz. Las algas fotosintéticas suelen tener un gran cloroplasto, pero las células vegetales tienen algunas decenas.
El medio interno del cloroplasto se llama estroma, y está compuesto por una disolución de glúcidos, lípidos, prótidos, ácidos nucleicos, pigmentos, nucleótidos, sales y elementos.
Su principal función es capturar la energía lumínica y utilizarla en la síntesis de materia orgánica.
Estructura de los cloroplastos
Los cloroplastos están formados por una doble membrana (externa e interna), un espacio intermembranoso y un espacio interior o estroma, donde se encuentran los tilacoides, con forma de sáculos aplanados. En el cloroplasto se distingue:
Membrana externa e interna. Su estructura es similar a la del resto de las membranas (60 por 100 son lípidos y el 40 por 100 proteínas). La membrana externa contiene porinas, por lo que es muy permeable, y la membrana interna, que es menos permeable, presenta proteínas de transporte específicas que regulan el paso de sustancias entre el hialoplasma y el estroma. Carecen de clorofila y, como en las mitocondrias, estas membranas tampoco tienen colesterol.
Espacio intermembrana. De composición muy parecido al citosol, por la permeabilidad de la membrana externa.
Tilacoides y grana. Son sáculos aplanados aislados o interconectados, parecidos a una pila de monedas formando una red interna membranosa. Se llama grana a cada uno de estos apilamientos, con un número variable de sacos. Las membranas de los tilacoides contienen todo lo necesario para realizar la fotosíntesis.
El 38 % son lípidos, el 50 % proteínas y el 12 % pigmentos (carotenoides y clorofilas).
De Kelvinsong - Trabajo propio, CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=23090800
De Kelvinsong - Trabajo propio, CC0, https://commons.wikimedia.org/w/index.php?curid=22952603
De Mnolf - Photo taken in Innsbruck, Austria, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=137710
By !Original: AgatellerVector: Qef - Own work, Public Domain, https://commons.wikimedia.org/w/index.php?curid=7072127
De Aibdescalzo - Own edit (Spanish translation) of File:Animal mitochondrion diagram en.svg by Mariana Ruiz (LadyofHats), Dominio público, https://commons.wikimedia.org/w/index.php?curid=6206970
El retículoendoplasmático es el orgánulo más grande de muchas células eucariotas. Está formado por túbulos (tubos) y sacos (cisternas) que se comunican formando una red continua desde la membrana nuclear, extendiéndose por todo el citoplasma de la célula. La parte interna del retículo endoplasmático se llama lumen(o espacio luminar o cisternal).
La membrana del RE tiene estructura similar a la plasmática, aunque es algo más delgada y tiene menos lípidos y más proteínas que la membrana plasmática.
Según sus funciones y su composición se distinguen dos tipos de retículo endoplasmático:
Retículo endoplasmático rugoso (RER)
Tiene ribosomas adheridos en sus membranas, en el lado del citosol. Los ribosomas están unidos por su subunidad mayor por la ayuda de unas proteínas del grupo de las riboforinas, que no se encuentran en el REL.
Se comunica con la membrana nuclear y el retículo endoplasmático liso.
Está formado por sacos y grandes cisternas aplanadas.
Su función principal es la de síntesis de proteínas con los ribosomas fijados a su membrana. Las proteínas sintetizadas pasan del RER al REL, luego al aparato de Golgi y de allí, a los lisosomas, a la membrana plasmática o al exterior. En el recorrido por estos orgánulos, la molécula experimentará un procesado adicional.
Las proteínas sintetizadas y almacenadas en el RER, antes de ser transportadas a otros orgánulos citoplasmáticos (aparato de Golgi, lisosomas), a la membrana plasmática o al exterior de la célula, deben ser glucosiladas para convertirse en glucoproteínas.La glucosilación de las proteínas se produce en el lumen del retículo, mientras que las proteínas del citosol no suelen estar glucosiladas.
Retículo endoplasmático liso (REL)
Carece de ribosomas, por lo que su superficie es lisa.
Formado, mayoritariamente, por una red de túbulos, que se unen al RER, que se extiende por todo el citoplasma.
La mayor parte de las células tienen un retículo endoplasmático liso escaso, pero es especialmente abundante en:
Células musculares estriadas.
Células intersticiales ováricas de Leydig, del testículo y células de la corteza suprarrenal que segregan hormonas esteroideas.
Hepatocitos, donde participa en la síntesis de moléculas lipoproteicas.
Funciones del retículo endoplasmático liso:
Síntesis, almacenamiento y transporte de lípidos.
En el lado citoplasmático de su membrana se sintetizan casi todos los lípidos constituyentes de las membranas: colesterol, fosfolípidos, glucolípidos, etc.
Los ácidos grasos se sintetizan en el citosol y entran en la membrana por acción de una flipasa.
En algunas células, el retículo endoplasmático liso produce grandes cantidades de lípidos, como por ejemplo, las hormonas esteroideas.
Contracción muscular. En las células del tejido muscular estriado, el REL libera calcio activando la contracción muscular.
Detoxificación. Su membrana contiene enzimas desintoxicantes que degradan sustancias liposolubles que puedan resultar nocivas y las transforman en sustancias solubles que pueden ser excretadas por el organismo. En los vertebrados, la detoxificación se realiza principalmente en las células del hígado, riñones, intestino, pulmones y piel. Por ejemplo, el REL abunda en las células del hígado, donde contiene enzimas que desintoxican productos nocivos como el alcohol y otros subproductos del metabolismo, como el amoníaco.
Liberación de glucosa a partir del glucógeno (en hepatocitos). El glucógeno almacenado en el hígado se encuentra en forma de pequeños gránulos adheridos a las membranas del REL. Cuando se necesita energía, el glucógeno se degrada obteniéndose glucosa-6-fosfato en el citoplasma. El REL elimina el grupo fosfato y las moléculas de glucosa entran en el REL y, son enviadas a la sangre hacia donde se requieran.
Produce vesículas de transporte con proteínas y lípidos recién sintetizados para llevarlos hacia el aparato de Golgi.
Aparato de Golgi
Aparato de Golgi con sus sectores
El aparato de Golgi es un orgánulo presente en todas las células eucariotas, excepto en los glóbulos rojos. Pertenece al sistema de endomembranas. Está formado por unas estructuras llamadas dictiosomas (dependiendo del tipo de célula), y estos dictiosomas están compuestos por unas 4 - 8cisternas (sáculos discoidales) aplanadas rodeadas de membrana que se encuentran apiladas unos encima de otras, y cuya función es completar la fabricación de algunas proteínas. Funciona como una planta empaquetadora, modificando vesículas del retículo endoplasmático rugoso. El material nuevo de las membranas se forma en varias cisternas del aparato de Golgi
Las células eucarióticas poseen un aparato de Golgi más o menos desarrollado, según la función que desempeñen. Los dictiosomas pueden presentar continuidad con otros componentes del sistema de endomembranas, como por ejemplo el retículo endoplasmático.
Estructura y composición del aparato de Golgi
Los dictiosomas presentan:
Una cara CIS, proximal o de formación: es la más interna, cercana al núcleo, y continuación de las membranas del retículo endoplasmático, del que recibe las vesículas de transición, que son sáculos con proteínas que han sido sintetizadas en la membrana del retículo endoplasmático rugoso (RER), introducidas dentro de sus cavidades y transportadas por el lumen hasta la parte más externa del retículo. El contenido de las vesículas de transición se incorpora a las cisternas del aparato de Golgi. De cada cisterna salen nuevas vesículas que se incorporan a la siguiente cisterna.
Su forma es convexa. Sus membranas son finas, de composición similar a la de las membranas del retículo endoplasmático.
Una cara intermedia o sáculos de la zona central, que tienen la mayor actividad metabólica. Presenta las vesículas intermedias o intercisternas.
Una cara TRANS, distal o de maduración: se encuentra próxima a la membrana plasmática (se parece más a ella). Su forma es cóncava. En la periferia de esta cara, se encuentran unas vesículas grandes llamadas vesículas de secreción, que contienen en su interior los productos finales y que llevarán a la membrana plasmática para expulsarlos al medio externo (exocitosis) o para formar lisosomas, que contienen enzimas digestivas.
Funciones del aparato de Golgi
Transporte y concentración de proteínas(formadas principalmente en el RE). Las proteínas procedentes del RER, englobadas en vesículas se unen a la región cis del dictiosoma. Aquí, si no están fosforiladas, las proteínas sufren una fosforilación. Las proteínas secretadas se van desplazando, pasando de una cisterna a otra, hasta llegar a las cisternas situadas en la cara trans del dictiosoma.
Glucosilación de lípidos y proteínas. Se completa la glicosilación que había comenzado en el RE. En el aparato de Golgi se produce el ensamblaje de oligosacáridos a lípidos y proteínas para formar glucolípidos y glucoproteínas respectivamente. También se fabrican los glucosaminoglucanos de la matriz extracelular de las células animales, así como las pectinas y la hemicelulosa de las paredes de las células vegetales.
Formación del del fragmoplasto en células vegetales. El tabique telofásico que divide el citoplasma en la zona ecuatorial de la célula vegetal, se produce por la asociación de vesículas derivadas del aparato de Golgi.
Formación del acrosoma en el espermatozoide. El acrosoma deriva del aparato de Golgi. Contiene enzimas hidrolíticas que sirven para digerir los componentes de las cubiertas del óvulo durante la fecundación.
Formación de membranas (cuando se expulsa al exterior el contenido de las vesículas mediante exocitosis, se unen a la membrana plasmática, incrementando la superficie).
Formación de lisosomas.
Concentración de sustancias (glúcidos, proteínas). Recolector y transporte.
Cuando se han sintetizado las glucoproteínas y los glucolípidos, irán a la membrana plasmática a través de las membranas de secreción.
Las proteínas (formadas en el RER) y los lípidos (formados en el REL) son transportados al aparato de Golgi por las vesículas de transición y allí van pasando de sáculo en sáculo hasta salir por las vesículas de secreción transformados en glucolípidos y glucoproteínas.
Los dictiosomas son estructuras muy dinámicas, ya que los sáculos del aparato de Golgi se renuevan constantemente por fusión de las vesículas de transición procedentes de las membranas del RE, y se van transformando los sáculos hasta llegar a ser vesículas de secreción.
Lisosomas
Los lisosomas son orgánulos globulares, formados por el aparato de Golgi, presentes tanto en células animales como vegetales.
Los lisosomas son vesículas membranosas, en cuyo interior tiene lugar la digestión intracelular. Su membrana es de tipo unitaria de 75 Å, recubierta por el interior por una gruesa capa de glucoproteínas que la protegen de la destrucción por la acción de las enzimas hidrolasas. La membrana del lisosoma contiene una bomba de protones que introduce H%2b y mantiene el pH ácido, inferior a 5, en el interior del lisosoma, y proteínas de transporte que permiten que pasen los productos resultantes de la digestión al citosol.
El lisosoma es el marcado con el número 12
Composición química y tipos de lisosomas
Los lisosomas contienen unas 50 enzimas diferentes del tipo hidrolasas ácidas, sintetizadas en el retículo endoplasmático rugoso. Son enzimas capaces de digerir las macromoléculas y tiene su funcionamiento óptimo en un medio con un pH = 5. La membrana del lisosoma contiene proteínas ácidas integrales altamente glucosiladas, que la protegen de las enzimas que contiene.
Los lisosomas contienen una gran cantidad de enzimas hidrolíticas, pero su contenido puede ser muy distinto. Se distinguen dos tipos de lisosomas:
Lisosomas primarios (también llamados “vesículas con hidrolasas”). Sólo contienen enzimas hidrolíticos. Son vesículas de secreción recién formadas por gemación a partir de las cisternas del aparato de Golgi. Pueden formar lisosomas secundarios o expulsar su contenido al exterior.
Lisosomas secundarios. También contienen hidrolasas, pero además sustratos en vía de digestión. Son lisosomas primarios fusionados con otras sustancias, de origen interno o externo. Según la sustancia que hidrolizan, se distinguen dos tipos:
Vacuolas heterofágicas: fusión de un lisosoma primario con partículas que proceden del exterior de la célula (fagolisosoma).
Vacuolas autofágicas: fusión de un lisosoma primario con distintas partes de la célula que ya no son necesarias o zonas lesionadas (autofagolisosoma).
Funciones de los lisosomas
Los lisosomas son los encargados de la digestión de las macromoléculas, aunque en algunos casos, sirven como almacén temporal de reservas:
1.- La digestión celular. Los lisosomas pueden digerir macromoléculas, tanto de origen interno como externo. Esta función la realizan de dos formas:
Autofagia. La materia que digiere es una parte de la célula, como fragmentos del RE, sáculos o cisternas del aparato de Golgi. La autofagia es importante para las células, ya que destruye zonas dañadas, interviene en el desarrollo celular y permite la nutrición de la célula cuando las condiciones son desfavorables.
Heterofagia. Lo digerido tiene origen externo. Tiene la doble finalidad de nutrir y defender la célula. Los sustratos entran a la célula por endocitosis, formando una vesícula que se fusiona a un lisosoma primario, originando a una vacuola digestiva, donde se produce la digestión. Los productos de la digestión pasan al hialoplasma, donde son reutilizados. Lo que no puede ser digerido, se expulsa al exterior por exocitosis, pudiendo fusionarse con la membrana plasmática.
2.- Almacenamiento de sustancias de reserva. En las semillas existe un tipo especial de lisosomas secundarios, los granos de aleurona, que almacenan las sustancias de reserva. No se produce la digestión celular hasta que no llega el momento de la germinación. Entonces se activan las enzimas hidrolíticas, iniciando la digestión intracelular, cuyos productos se utilizarán en el desarrollo del embrión.
Vacuolas
Vacuolas en rosa
Las vacuolas y las inclusiones son lugares del interior de la célula en los que se acumulan sustancias. Si están rodeados por una membrana se denominan vacuolas; en caso contrario, inclusiones.
Son orgánulos rodeados por una membrana simple, con forma redondeada y que contienen sustancias que no están involucradas en ningún proceso metabólico. Se forman en células jóvenes, por fusión de vesículas derivadas del retículo endoplasmático y del aparato de Golgi.
Las células eucariotas animales tienen bastantes vacuolas. Las células eucariotas vegetales, cuando son jóvenes tienen muchas vacuolas, pero cuando son maduras pueden llegar a tener una única célula que ocupa el 90% del volumen celular.
Dentro de la vacuola se encuentra el jugo vacuolar amorfo, formado por agua y que puede contener diversas sustancias de almacén o de desecho.
El conjunto de vacuolas de una célula recibe el nombre de vacuoma.
Tipos de vacuolas
Vacuola vegetal
Las células vegetales jóvenes tienen muchas vacuolas pequeñas porque contienen poca agua, pero según van creciendo, aumentan de tamaño y se fusionan formando una gran vacuola central que ocupa casi todo el citoplasma.
Las vacuolas vegetales son generalmente incoloras (excepto las de los pétalos de las flores). La membrana que formavacuola vegetal se llama tonoplasto.
La principal función de las vacuolas en vegetales consiste en almacenar sustancias.
Vacuola contráctil (vacuola pulsátil)
En las células animales, las vacuolas están relacionadas con el aparato de Golgi, con funciones de almacenamiento de sustancias, transporte, etc. Ya hemos hablado de ellas cuando vimos los lisosomas: vacuolas digestivas yvacuolas autofágicas.
Entre las vacuolas animales destacan las vacuolas pulsátiles que presentan las células que viven en ambientes hipotónicos, como es el caso de muchos protozoos, y que son utilizadas para regular la flotabilidad bombeando el exceso de agua al exterior por transporte activo.
Funciones de las vacuolas
Las funciones principales de las vacuolas son:
Mantenimiento de la turgencia celular. La presión osmótica dentro de las vacuolas es muy alta por la elevada concentración de sustancias. El agua tiende a penetrar en las vacuolas por ósmosis, lo que provoca que la célula se mantenga turgente, contribuyendo a que la planta esté erguida.
Permiten que aumente de tamaño la célula vegetal, sin que ello suponga un excesivo gasto de energía por su parte.
Almacenamiento de sustancias diversas. Las vacuolas pueden servir como almacén para muchas moléculas de la célula:
Agua.
Sustancias de reserva, como glúcidos, ácidos grasos y proteínas.
Productos de desecho.
Pigmentos, como antocianinas y flavonas de los pétalos de las flores.
Sustancias de desecho resultantes del metabolismo celular y que pueden resultar tóxicas para la planta si están libres en el citoplasma. Por ejemplo, la nicotina o el cianuro.
Perixosomas
Estructura básica del perixosoma
Los peroxisomas son unos orgánulos esféricos, pequeños, presentes en células animales y vegetales, rodeados de una membrana unitaria de 75Å, que contienen unas enzimas oxidasas, entre las que destacan la peroxidasa y la catalasa.
Los peroxisomas son unos orgánulos muy parecidos a los lisosomas, pero se diferencian de ellos en que no contienen hidrolasas ácidas, sino enzimas oxidativos.
Son abundantes en células que sintetizan, almacenan o descomponen lípidos.
En los peroxisomas se realizan reacciones de oxidación similares a las producidas en las mitocondrias, pero la energía producida se disipa en forma de calor, en lugar de almacenarse en forma de ATP.
La peroxidasa utiliza oxígeno molecular para oxidar diversos tipos de sustratos desprendiendo peróxido de hidrógeno (H2O2), sustancia que es tóxica para la célula. Después actúa la catalasa, que descompone el peróxido de hidrógeno.
Así, pueden oxidar ácidos grasos y aminoácidos, aportando energía para la célula, además de detoxificar una gran variedad de moléculas tóxicas, sobre todo en el hígado y en el riñón.
Las funciones de los peroxisomas son:
Detoxificación de algunas sustancias tóxicas para el organismo, como ácido úrico, etanol, metanol, …
Degradación de ácidos grasos y aminoácidos que no van a generar ATP, como ocurre en la mitocondria, sino energía en forma de calor.
Gluxisomas
Un tipo especial de peroxisomas son los llamados glioxisomas, exclusivos de células vegetales, que aparecen en semillas en germinación y permiten producir glúcidos a partir de los ácidos grasos.
Mitocondias
Las mitocondrias están presentes en casi todas las células eucarióticas, mientras que los cloroplastos sólo aparecen en las células vegetales fotosintéticas. Los dos se incluyen dentro de los orgánulos energéticosde las células.
Si no fuera por las mitocondrias, las células heterótrofas sólo podrían obtener su ATP de la glucólisis anaerobia, pero con ellas pueden degradar totalmente la glucosa en presencia de oxígeno hasta convertirla en dióxido de carbono y agua, obteniendo mucha más energía.
El número de mitocondrias depende de las necesidades energéticas de la célula, situándose en zonas en las que se requiere un consumo importante de ATP.
Estructura de la mitocondrias
Las mitocondrias tienen forma cilíndrica y están limitadas por una doble membrana, la membrana mitocondrial externa, que la separa del hialoplasma, y la membrana mitocondrial interna, que tienen unos repliegues hacia el interior que aumentan su superficie, las crestas mitocondriales. Estas membranas dejan dos compartimentos: el espacio intermembrana, entre ambas membranas, y la matriz mitocondrial, espacio que rodea la membrana mitocondrial interna.
Funciones de las mitocondrias
La función principal de las mitocondrias es la obtención de energía, aunque cada componente mitocondrial tiene distinta función:
El hecho de que los compartimentos mitocondriales tengan distintos complejos enzimáticos, implica que en cada uno de ellos se realizarán funciones diferentes:
Ciclo de Krebs. Tiene lugar en la matriz mitocondrial.
Cadena respiratoria. En el ciclo de Krebs se desprenden una serie de electrones. Los transportadores de electrones se localizan en la membrana mitocondrial interna.
Fosforilación oxidativa. Se realiza en las crestas mitocondriales. La ATP-asa fosforila el ADP y lo transforma en ATP. Así se sintetiza la mayor parte del ATP que se produce en las células aerobias.
La β-oxidación de los ácidos grasos. Las enzimas que intervienen en ella están situadas en la matriz mitocondrial.
Concentración de sustancias en la cámara interna, tales como proteínas, lípidos, colorantes, hierro, plata, calcio, fosfatos y partículas semejantes a los virus.
Cloroplasto
Los cloroplastos son un tipo de cromoplastos que se localizan en las células vegetales fotosintéticas y en las algas verdes.
Se pueden mover por movimientos ameboideos y contráctiles, y se colocan en la parte de la célula que recibe mayor cantidad de luz. Las algas fotosintéticas suelen tener un gran cloroplasto, pero las células vegetales tienen algunas decenas.
El medio interno del cloroplasto se llama estroma, y está compuesto por una disolución de glúcidos, lípidos, prótidos, ácidos nucleicos, pigmentos, nucleótidos, sales y elementos.
Su principal función es capturar la energía lumínica y utilizarla en la síntesis de materia orgánica.
Estructura de los cloroplastos
Los cloroplastos están formados por una doble membrana (externa e interna), un espacio intermembranoso y un espacio interior o estroma, donde se encuentran los tilacoides, con forma de sáculos aplanados. En el cloroplasto se distingue:
Membrana externa e interna. Su estructura es similar a la del resto de las membranas (60 por 100 son lípidos y el 40 por 100 proteínas). La membrana externa contiene porinas, por lo que es muy permeable, y la membrana interna, que es menos permeable, presenta proteínas de transporte específicas que regulan el paso de sustancias entre el hialoplasma y el estroma. Carecen de clorofila y, como en las mitocondrias, estas membranas tampoco tienen colesterol.
Espacio intermembrana. De composición muy parecido al citosol, por la permeabilidad de la membrana externa.
Tilacoides y grana. Son sáculos aplanados aislados o interconectados, parecidos a una pila de monedas formando una red interna membranosa. Se llama grana a cada uno de estos apilamientos, con un número variable de sacos. Las membranas de los tilacoides contienen todo lo necesario para realizar la fotosíntesis.
El 38 % son lípidos, el 50 % proteínas y el 12 % pigmentos (carotenoides y clorofilas).
De Kelvinsong - Trabajo propio, CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=23090800
De Kelvinsong - Trabajo propio, CC0, https://commons.wikimedia.org/w/index.php?curid=22952603
De Mnolf - Photo taken in Innsbruck, Austria, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=137710
By !Original: AgatellerVector: Qef - Own work, Public Domain, https://commons.wikimedia.org/w/index.php?curid=7072127
De Aibdescalzo - Own edit (Spanish translation) of File:Animal mitochondrion diagram en.svg by Mariana Ruiz (LadyofHats), Dominio público, https://commons.wikimedia.org/w/index.php?curid=6206970
Juega a aprender el cuerpo humano, las células y química
Más de 200 juegos interactivos para aprender las células, animal y vegetal, así como los huesos, músculos, aparatos y sistemas del cuerpo humano, la tabla periódica, los estados de agregación de la materia y la estructura del átomo (modelo de Bohr).
Diversos tipos de juego con analíticas, retos, torneos y ranking.
12 cuadernos de cálculo con ejercicios, problemas y actividades de refuerzo para afianzar el conocimiento de las sumas, restas, multiplicaciones y divisiones.
También incluyen problemas para cuya resolución se debe aplicar alguna o varias de las operaciones matemáticas básicas.
¡Recortables de papel que representan a los personajes más famosos de la historia!
32 Paper Toys que introducen a los niños en las profesiones así como en la historia del arte, la ciencia y la literatura por medio de algunos de sus protagonistas más destacados.
Juega a aprender geografía con los mapas más utilizados en los coles
Un pack de juegos interactivos de geografía de todo el mundo:
· Más de 500 juegos inteligentes
· Retos educativos entre alumnos, juego uno contra uno
· Torneos, el reto se hace colectivo con desafíos entre grupos de jugadores
· Diversos niveles de dificultad y modalidades de juego
· Modo estudia para memorizar antes de consolidar lo aprendido en el juego
· Analíticas de juego, datos y estadísticas pertinentes para medir los impactos del aprendizaje
· Recomendaciones de juegos a practicar y personas a quienes retar
· Ranking de juegos para cada juego individual, por ámbitos (geografía de Europa, América,...) y ranking general de Geografía
· Multiidoma, todos los juegos en 9 idiomas diferentes
· Accesibles desde cualquier dispositivo: móvil, tablet u ordenador
Más información
Reta a tus amigos a jugar a anatomía humana y celular y química de manera divertida y eficaz
Un pack de juegos interactivos de anatomía humana y celula y químicar:
· Más de 200 juegos inteligentes
· Retos educativos entre alumnos, juego uno contra uno
· Torneos, el reto se hace colectivo con desafíos entre grupos de jugadores
· Diversos niveles de dificultad y modalidades de juego
· Modo estudia para memorizar antes de consolidar lo aprendido en el juego
· Analíticas de juego, datos y estadísticas pertinentes para medir los impactos del aprendizaje
· Recomendaciones de juegos a practicar y personas a quienes retar
· Ranking de juegos para cada juego individual, por ámbitos anatómicos (sistema esquelético, muscular, anatomía celular...), por química y ranking general de anatomía y química
· Multiidoma, todos los juegos en 9 idiomas diferentes
· Accesibles desde cualquier dispositivo: móvil, tablet u ordenador
Más información
Ejercita la caligrafía y mejora en escritura y ortografía
La colección Cuadernos de Caligrafía de Didactalia está pensada para que los alumnos de primaria ejerciten y dominen la caligrafía siguiendo las pautas Montessori ydoble línea. Incluye 16 cuadernos de caligrafía con diversos ejercicios de copia de letras, sílabas, palabras y frases, así como actividades relacionadas con las normas de ortografía.
Cada cuaderno se ajusta a un nivel educativo, ampliando de
forma progresiva su nivel de dificultad.
Más información
Domina el cálculo y el manejo de los números
La colección Cuadernos de Cálculo de Didactalia está formada por 12 cuadernos de cálculo, también para primaria. Su
objetivo es lograr que los alumnos dominen el cálculo y el manejo de los números mediante la práctica sistemática de
las cuatro operaciones matemáticas básicas: sumas, restas, multiplicaciones y divisiones.
Cada cuaderno se ajusta a un nivel educativo, ampliando de forma progresiva su nivel de dificultad.
Más información
Conoce a los personajes más famosos de la historia
32 recortables de papel que presentan a los más pequeños, de una forma divertida, a algunos de los personajes más famosos
de la historia, así como diferentes oficios y profesiones. Los recortables facilitan además el desarrollo de la motricidad, esencial en el desarrollo infantil.
Goya, Da Vinci o Cervantes, son algunas de las figuras destacadas de las artes, las ciencias o las letras incluidas en la colección de Papertoys de Grandes Personajes, pensada para que los más pequeños puedan acercarse a estos célebres personajes y sus obras.
Más información
Aprende historia de una manera diferente y divertida
Un pack de juegos interactivos de historia:
· Más de 800 juegos inteligentes
· Retos educativos entre alumnos, juego uno contra uno
· Torneos, el reto se hace colectivo con desafíos entre grupos de jugadores
· Diversos niveles de dificultad y modalidades de juego
· Modo estudia para memorizar antes de consolidar lo aprendido en el juego
· Analíticas de juego, datos y estadísticas pertinentes para medir los impactos del aprendizaje
· Recomendaciones de juegos a practicar y personas a quienes retar
· Ranking de juegos para cada juego individual y ranking general de Timelines
· Multiidoma, todos los juegos en 9 idiomas diferentes
· Accesibles desde cualquier dispositivo: móvil, tablet u ordenador






































Comment
0