
Cargando...
Que pode fazer?

Los glúcidos son biomoléculas formadas básicamente por carbono (C), hidrógeno (H) y oxígeno (O), en una proporción semejante a CnH2nOn, es decir (CH2O)n. También se les suele llamar hidratos de carbono o carbohidratos, aunque este nombre es poco apropiado, ya que no se trata de átomos de carbono hidratados, enlazados a moléculas de agua, sino de átomos de carbono unidos a grupos alcohol (-OH), llamados también radicales hidroxilo, y a radicales hidrógeno (-H). El nombre glúcido deriva de la palabra “glucosa”, del griego glykys, que significa “dulce”.
Los monosacáridos son glúcidos constituidos por una sola cadena polihidroxialdehídica (aldosa) o polihidroxicetónica (cetosas). No pueden descomponerse mediante hidrólisis. Se nombran añadiendo la terminación -osa al número de carbonos, como por ejemplo, triosa, tetrosa, pentosa, hexosa, etc.
Una cetosa es un monosacárido con un grupo cetona por molécula. La dihidroxiacetona, de tres átomos de carbono, es la cetosa más simple y la única que no presenta actividad óptica.
Por tanto, las aldosas y las cetosas, tienen un grupo carbonilo (carbono unido mediante doble enlace al oxígeno).
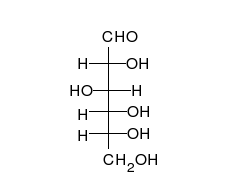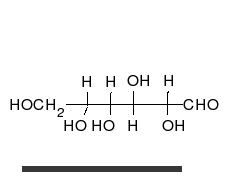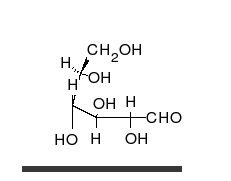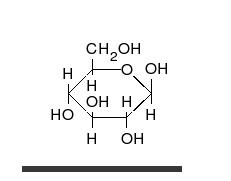
Los monosacáridos pueden ser oxidados (o ellos reducir) por determinados iones como el ion férrico (Fe3+) y cúprico (Cu2+) que pasarán al reducirse a Fe2+ o Cu+1. El grupo carbonilo de los monosacáridos pasa a ácido cuando se oxida.
Ésta es la base de la reacción de Fehling en la que una disolución azul, por la presencia de Cu2+, pasa a Cu+, de color rojo, en presencia de azúcares reductores como la glucosa.
Otra propiedad química de los glúcidos es su capacidad para asociarse con grupos amino -NH2.
Hay dos tipos de enlaces entre un monosacárido y otras moléculas:
El enlace O-glucosídico es α-glucosídico si el primer monosacárido es α, y β-glucosídico si el primer monosacárido es β. Por ejemplo, entre el C1 de una α-D-glucopiranosa y el C4 de otra D-glucopiranosa (α o β) se establece un enlace tipo α (1→4).
En el enlace O-glucosídico reacciona el grupo OH (hidroxilo) del carbono anomérico del primer monosácarido con un OH unido a un carbono (anomérico o no) del segundo monosacárido. Se forma un disacárido y una molécula de agua. El proceso es realmente una condensación, se denomina deshidratación por la característica de la pérdida de la molécula de agua, al igual que ocurre en la formación del enlace peptídico.
Si los OH de la reacción provienen de los dos carbonos anómericos, el disacárido será dicarbonílico y no tendrá poder reductor. Sin embargo, si en el enlace participan los OH de un carbono anomérico y de otro carbono no anomérico, el disacárido será monocarbonílico y tendrá poder reductor (ya que queda un grupo OH libre en el otro carbono anómerico). Este hecho se puede comprobar exprimentalmente mediante la reacción con el reactivo de Fehling o con el reactivo de Tollens.
Al final del proceso ambos monosacáridos quedarán unidos por un oxígeno (O).
Para romper los enlaces O-glucosídicos será necesaria la hidrólisis, añadiendo una molécula de agua.
Los enlaces glucosídicos se distinguen por el tipo de anómero de los monosacáridos que intervienen y de los carbonos que intervienen en el enlace. Así hay enlaces α(1→6), β(1→4), etc.

Los disacáridos están formados por la unión de dos monosacáridos mediante un enlace O-glucosídico, que se puede realizarse de dos formas:
Los disacáridos presentan las mismas propiedades que los monosacáridos: son solubles en agua, cristalizables, incoloros y de sabor dulce. Su capacidad reductora depende de si tiene un grupo anomérico libre.
Los principales disacáridos con interés biológico son:
Disacárido formado por dos moléculas de D-glucopiranosa unidas mediante enlace α (1→4), entre el oxígeno del primer carbono anomérico (proveniente de -OH) de una glucosa y el oxígeno perteneciente al cuarto carbono de la otra. La maltosa es el α -D-glucopiranosil (1 →4)-D-glucopiranosa y al tener libre el carbono anomérico del segundo es un disacárido reductor.
A la maltosa se le llama también azúcar de malta, ya que aparece en los granos germinados de cebada.
Disacárido formado por dos moléculas de D-glucopiranosa unidas mediante enlace β (1→4). No se encuentra libre en la naturaleza. Se obtiene por hidrólisis de la celulosa.
La lactosa es un disacárido formado por la unión de una molécula de D-galactopiranosa y otra de D-glucopiranosa unidas por medio de un enlace β (1→4). Se encuentra libre en la leche de los mamíferos.
Disacárido formado por una molécula de α-D-glucopiranosa y otra de β-D-fructofuranosa unidas por medio de un enlace α (1→2). El enlace se realiza entre el -OH del carbono anomérico del primer monosacárido y el -OH del carbono anomérico del segundo. Debido a este enlace dicarbonílico, es el único disacárido de los citados que no tiene poder reductor sobre el licor de Fehling.
Se encuentra en la caña de azúcar y en la remolacha azucarera, es el azúcar de consumo habitual.
Disacárido formado por dos moléculas de D-glucopiranosa unidas por enlace α (1→6). No se encuentra libre en la naturaleza. Se obtiene por hidrólisis de la amilopectina (un componente del almidón y del glucógeno). Proviene de los puntos de ramificación α de estos polisacáridos.

Los polisacáridos son biomoléculas formadas por la unión de muchos monosacáridos (incluso miles) mediante enlaces O-glucosídicos, desprendiéndose una molécula de agua por cada enlace.
Sus pesos moleculares son muy altos. No tienen sabor dulce. Pueden ser insolubles, como la celulosa, o formar dispersiones coloidales, como el almidón. No tienen poder reductor.
Pueden desempeñar funciones estructurales o de reserva energética. Los polisacáridos que realizan una función estructural presentan enlace β-glucosídico, y los que llevan a cabo una función de reserva energética presentan enlace α-glucosídico.
Distinguiremos:
Los homopolisacáridos se clasifican según el tipo de monosacárido que se repite y según el tipo de enlace.
Su función depende del tipo de enlace O-glucosídico.
Homopolisacáridos de reserva:
El almidón es el polisacárido de reserva propio de los vegetales. Se acumula en forma de gránulos dentro de los plastos, en la célula vegetal.
El almidón está formado por miles de moléculas de glucosa. Como no están disueltas en el citoplasma, no influyen en la presión osmótica interna y constituyen una gran reserva energética que ocupa poco volumen.
El almidón se encuentra en las semillas y en los tubérculos, como la patata y el boniato. Con esta reserva energética, las plantas pueden obtener energía sin necesidad de luz.
Pero el almidón no es realmente un polisacárido, sino la mezcla de dos, la amilosa (30 %) y la amilopectina (70 %).
Los animales pueden digerir el almidón (no la celulosa) mediante la acción de varias enzimas:
El glucógeno es el polisacárido de reserva energética propio de los animales y hongos (y en algunas bacterias). Se encuentra en el hígado y en los músculos.
El glucógeno, al igual que la amilopectina, está constituido por un polímero de glucosas (α-D-glucopiranosa) unidas mediante enlaces α (1→4), con ramificaciones en posición α (1→6). Su estructura es semejante a la de la amilopectina, aunque con más ramificaciones, cada ocho o diez glucosas.
Con el yodo, la dispersión coloidal se tiñe de rojo oscuro. Las enzimas amilasas sobre el glucógeno dan maltosas y dextrina límite. Luego, mediante las enzimas R-desramificantes y las maltasas, se obtiene glucosa.
Son los polisacáridos de reserva de las levaduras. Están compuestos por cadenas ramificadas de α-D-glucopiranosas, pero las uniones son variadas: (1→2), (1→3), etcétera. La placa dental bacteriana es rica en dextranos.

La celulosa es un polisacárido con función esquelética propio de los vegetales. Es el elemento principal de la pared celular, que envuelve a la célula, y persiste después de la muerte de ésta. Las fibras vegetales (algodón, lino, cáñamo, esparto, etc.) y el interior del tronco de los árboles (el leño o madera) están formados, principalmente, por paredes celulósicas de células muertas.
La celulosa es un polímero de β-D-glucopiranosas unidas mediante enlaces β (1→4). Dos glucosas forman una celobiosa. Cada polímero tiene de 150 a 5000 moléculas de celobiosas, y un peso molecular medio de 800.000. Estas cadenas de polímeros no están ramificadas, pero se pueden disponer paralelamente uniéndose mediante enlaces de puente de hidrógeno, en agregados cristalinos muy ordenados.
Los humanos no podemos digerir la celulosa porque nuestras enzimas digestivas no pueden romper el enlace β. Muchos microorganismos y algunos invertebrados (Lepisma saccharina o pececillo de plata, y el Teredo navalis o molusco taladrador de la madera) son capaces de segregar la enzima celulasa, que puede romper el enlace β. Los insectos xilófagos, como los termes, y los herbívoros rumiantes (vaca, oveja, cabra, camello), pueden aprovechar la celulosa porque los microorganismos simbióticos del tracto digestivo producen celulasa. Los herbívoros no rumiantes, como el caballo, presentan restos de celulosa en las heces.
La celulosa es un nutriente importante para muchos animales, aunque la mayoría de las especies requieren la ayuda de los organismos simbióticos para que produzcan celulasa, la enzima capaz de romper el enlace β glucosídico.
Es un polisacárido estructural, componente fundamental del exoesqueleto de los artrópodos (insectos, crustáceos, etc.). También forma parte de los recubrimientos celulares de los hongos. El monómero constituyente es un derivado de la glucosa (N-acetil-β-D-glucosamina). La unión entre ellos se realiza por enlaces β (1→4), que da lugar a una cadena lineal, similar a la celulosa. Tampoco es digerible por los animales.
Los heteropolisacáridos son sustancias que, por hidrólisis, dan lugar a varios tipos distintos de monosacáridos o de derivados de éstos. Los principales son:
Los heterósidos se forma por la unión de un glúcido con otra molécula no glucídica, que denominamos aglucón.
Los principales tipos de asociación entre glúcidos y otros tipos de moléculas son:
Los peptidoglicanos o mureína son polímeros de N-acetilglucosamina (NAG) y N-acetilmurámico (NAM) unidos mediante enlaces β(1→4), unidas a cadenas de aminoácidos cortas. Forman parte de la pared bacteriana.
Los proteoglicanos son moléculas formadas por una gran fracción de polisacáridos (80 % de la molécula), denominados glucosaminglucanos, y una pequeña fracción proteica (aproximadamente 20 %).
Las cadenas de glucosaminoglucanos están formadas por polímeros lineales de N–acetilglucosamina (o N-acetilgalactosamina) y ácido glucurónico. Los más habituales son:
Las glicoproteínas son moléculas formadas por una pequeña fracción glucídica (entre un 5 % y un 40 %) y una gran fracción proteica, que se unen mediante enlaces fuertes (covalentes).
Se diferencian de los proteoglucanos en que la parte glucídica no contiene ni ácido hialurónico ni sulfatos de condroitina.
Las principales son: las mucinas de secreción, como las salivales; las glucoproteínas de la sangre, como la protrombina y las inmunoglobulinas; las hormonas gonadotrópicas; algunas enzimas ribonucleasas y las denominadas glicoproteínas de las membranas celulares.
Los glucolípidos están constituidos por monosacáridos u oligosacáridos unidos a lípidos (ceramida). Generalmente se encuentran en la membrana celular. Los más conocidos son:
Texto:
Imagen:
Los glúcidos son biomoléculas formadas básicamente por carbono (C), hidrógeno (H) y oxígeno (O), en una proporción semejante a CnH2nOn, es decir (CH2O)n. También se les suele llamar hidratos de carbono o carbohidratos, aunque este nombre es poco apropiado, ya que no se trata de átomos de carbono hidratados, enlazados a moléculas de agua, sino de átomos de carbono unidos a grupos alcohol (-OH), llamados también radicales hidroxilo, y a radicales hidrógeno (-H). El nombre glúcido deriva de la palabra “glucosa”, del griego glykys, que significa “dulce”.
Los monosacáridos son glúcidos constituidos por una sola cadena polihidroxialdehídica (aldosa) o polihidroxicetónica (cetosas). No pueden descomponerse mediante hidrólisis. Se nombran añadiendo la terminación -osa al número de carbonos, como por ejemplo, triosa, tetrosa, pentosa, hexosa, etc.
Una cetosa es un monosacárido con un grupo cetona por molécula. La dihidroxiacetona, de tres átomos de carbono, es la cetosa más simple y la única que no presenta actividad óptica.
Por tanto, las aldosas y las cetosas, tienen un grupo carbonilo (carbono unido mediante doble enlace al oxígeno).
Los monosacáridos pueden ser oxidados (o ellos reducir) por determinados iones como el ion férrico (Fe3%2b) y cúprico (Cu2%2b) que pasarán al reducirse a Fe2%2b o Cu%2b1. El grupo carbonilo de los monosacáridos pasa a ácido cuando se oxida.
Ésta es la base de la reacción de Fehling en la que una disolución azul, por la presencia de Cu2%2b, pasa a Cu%2b, de color rojo, en presencia de azúcares reductores como la glucosa.
Otra propiedad química de los glúcidos es su capacidad para asociarse con grupos amino -NH2.
Hay dos tipos de enlaces entre un monosacárido y otras moléculas:
El enlace O-glucosídico es α-glucosídico si el primer monosacárido es α, y β-glucosídico si el primer monosacárido es β. Por ejemplo, entre el C1 de una α-D-glucopiranosa y el C4 de otra D-glucopiranosa (α o β) se establece un enlace tipo α (1→4).
En el enlace O-glucosídico reacciona el grupo OH (hidroxilo) del carbono anomérico del primer monosácarido con un OH unido a un carbono (anomérico o no) del segundo monosacárido. Se forma un disacárido y una molécula de agua. El proceso es realmente una condensación, se denomina deshidratación por la característica de la pérdida de la molécula de agua, al igual que ocurre en la formación del enlace peptídico.
Si los OH de la reacción provienen de los dos carbonos anómericos, el disacárido será dicarbonílico y no tendrá poder reductor. Sin embargo, si en el enlace participan los OH de un carbono anomérico y de otro carbono no anomérico, el disacárido será monocarbonílico y tendrá poder reductor (ya que queda un grupo OH libre en el otro carbono anómerico). Este hecho se puede comprobar exprimentalmente mediante la reacción con el reactivo de Fehling o con el reactivo de Tollens.
Al final del proceso ambos monosacáridos quedarán unidos por un oxígeno (O).
Para romper los enlaces O-glucosídicos será necesaria la hidrólisis, añadiendo una molécula de agua.
Los enlaces glucosídicos se distinguen por el tipo de anómero de los monosacáridos que intervienen y de los carbonos que intervienen en el enlace. Así hay enlaces α(1→6), β(1→4), etc.
Los disacáridos están formados por la unión de dos monosacáridos mediante un enlace O-glucosídico, que se puede realizarse de dos formas:
Los disacáridos presentan las mismas propiedades que los monosacáridos: son solubles en agua, cristalizables, incoloros y de sabor dulce. Su capacidad reductora depende de si tiene un grupo anomérico libre.
Los principales disacáridos con interés biológico son:
Disacárido formado por dos moléculas de D-glucopiranosa unidas mediante enlace α (1→4), entre el oxígeno del primer carbono anomérico (proveniente de -OH) de una glucosa y el oxígeno perteneciente al cuarto carbono de la otra. La maltosa es el α -D-glucopiranosil (1 →4)-D-glucopiranosa y al tener libre el carbono anomérico del segundo es un disacárido reductor.
A la maltosa se le llama también azúcar de malta, ya que aparece en los granos germinados de cebada.
Disacárido formado por dos moléculas de D-glucopiranosa unidas mediante enlace β (1→4). No se encuentra libre en la naturaleza. Se obtiene por hidrólisis de la celulosa.
La lactosa es un disacárido formado por la unión de una molécula de D-galactopiranosa y otra de D-glucopiranosa unidas por medio de un enlace β (1→4). Se encuentra libre en la leche de los mamíferos.
Disacárido formado por una molécula de α-D-glucopiranosa y otra de β-D-fructofuranosa unidas por medio de un enlace α (1→2). El enlace se realiza entre el -OH del carbono anomérico del primer monosacárido y el -OH del carbono anomérico del segundo. Debido a este enlace dicarbonílico, es el único disacárido de los citados que no tiene poder reductor sobre el licor de Fehling.
Se encuentra en la caña de azúcar y en la remolacha azucarera, es el azúcar de consumo habitual.
Disacárido formado por dos moléculas de D-glucopiranosa unidas por enlace α (1→6). No se encuentra libre en la naturaleza. Se obtiene por hidrólisis de la amilopectina (un componente del almidón y del glucógeno). Proviene de los puntos de ramificación α de estos polisacáridos.
Los polisacáridos son biomoléculas formadas por la unión de muchos monosacáridos (incluso miles) mediante enlaces O-glucosídicos, desprendiéndose una molécula de agua por cada enlace.
Sus pesos moleculares son muy altos. No tienen sabor dulce. Pueden ser insolubles, como la celulosa, o formar dispersiones coloidales, como el almidón. No tienen poder reductor.
Pueden desempeñar funciones estructurales o de reserva energética. Los polisacáridos que realizan una función estructural presentan enlace β-glucosídico, y los que llevan a cabo una función de reserva energética presentan enlace α-glucosídico.
Distinguiremos:
Los homopolisacáridos se clasifican según el tipo de monosacárido que se repite y según el tipo de enlace.
Su función depende del tipo de enlace O-glucosídico.
Homopolisacáridos de reserva:
El almidón es el polisacárido de reserva propio de los vegetales. Se acumula en forma de gránulos dentro de los plastos, en la célula vegetal.
El almidón está formado por miles de moléculas de glucosa. Como no están disueltas en el citoplasma, no influyen en la presión osmótica interna y constituyen una gran reserva energética que ocupa poco volumen.
El almidón se encuentra en las semillas y en los tubérculos, como la patata y el boniato. Con esta reserva energética, las plantas pueden obtener energía sin necesidad de luz.
Pero el almidón no es realmente un polisacárido, sino la mezcla de dos, la amilosa (30 %) y la amilopectina (70 %).
Los animales pueden digerir el almidón (no la celulosa) mediante la acción de varias enzimas:
El glucógeno es el polisacárido de reserva energética propio de los animales y hongos (y en algunas bacterias). Se encuentra en el hígado y en los músculos.
El glucógeno, al igual que la amilopectina, está constituido por un polímero de glucosas (α-D-glucopiranosa) unidas mediante enlaces α (1→4), con ramificaciones en posición α (1→6). Su estructura es semejante a la de la amilopectina, aunque con más ramificaciones, cada ocho o diez glucosas.
Con el yodo, la dispersión coloidal se tiñe de rojo oscuro. Las enzimas amilasas sobre el glucógeno dan maltosas y dextrina límite. Luego, mediante las enzimas R-desramificantes y las maltasas, se obtiene glucosa.
Son los polisacáridos de reserva de las levaduras. Están compuestos por cadenas ramificadas de α-D-glucopiranosas, pero las uniones son variadas: (1→2), (1→3), etcétera. La placa dental bacteriana es rica en dextranos.
La celulosa es un polisacárido con función esquelética propio de los vegetales. Es el elemento principal de la pared celular, que envuelve a la célula, y persiste después de la muerte de ésta. Las fibras vegetales (algodón, lino, cáñamo, esparto, etc.) y el interior del tronco de los árboles (el leño o madera) están formados, principalmente, por paredes celulósicas de células muertas.
La celulosa es un polímero de β-D-glucopiranosas unidas mediante enlaces β (1→4). Dos glucosas forman una celobiosa. Cada polímero tiene de 150 a 5000 moléculas de celobiosas, y un peso molecular medio de 800.000. Estas cadenas de polímeros no están ramificadas, pero se pueden disponer paralelamente uniéndose mediante enlaces de puente de hidrógeno, en agregados cristalinos muy ordenados.
Los humanos no podemos digerir la celulosa porque nuestras enzimas digestivas no pueden romper el enlace β. Muchos microorganismos y algunos invertebrados (Lepisma saccharina o pececillo de plata, y el Teredo navalis o molusco taladrador de la madera) son capaces de segregar la enzima celulasa, que puede romper el enlace β. Los insectos xilófagos, como los termes, y los herbívoros rumiantes (vaca, oveja, cabra, camello), pueden aprovechar la celulosa porque los microorganismos simbióticos del tracto digestivo producen celulasa. Los herbívoros no rumiantes, como el caballo, presentan restos de celulosa en las heces.
La celulosa es un nutriente importante para muchos animales, aunque la mayoría de las especies requieren la ayuda de los organismos simbióticos para que produzcan celulasa, la enzima capaz de romper el enlace β glucosídico.
Es un polisacárido estructural, componente fundamental del exoesqueleto de los artrópodos (insectos, crustáceos, etc.). También forma parte de los recubrimientos celulares de los hongos. El monómero constituyente es un derivado de la glucosa (N-acetil-β-D-glucosamina). La unión entre ellos se realiza por enlaces β (1→4), que da lugar a una cadena lineal, similar a la celulosa. Tampoco es digerible por los animales.
Los heteropolisacáridos son sustancias que, por hidrólisis, dan lugar a varios tipos distintos de monosacáridos o de derivados de éstos. Los principales son:
Los heterósidos se forma por la unión de un glúcido con otra molécula no glucídica, que denominamos aglucón.
Los principales tipos de asociación entre glúcidos y otros tipos de moléculas son:
Los peptidoglicanos o mureína son polímeros de N-acetilglucosamina (NAG) y N-acetilmurámico (NAM) unidos mediante enlaces β(1→4), unidas a cadenas de aminoácidos cortas. Forman parte de la pared bacteriana.
Los proteoglicanos son moléculas formadas por una gran fracción de polisacáridos (80 % de la molécula), denominados glucosaminglucanos, y una pequeña fracción proteica (aproximadamente 20 %).
Las cadenas de glucosaminoglucanos están formadas por polímeros lineales de N–acetilglucosamina (o N-acetilgalactosamina) y ácido glucurónico. Los más habituales son:
Las glicoproteínas son moléculas formadas por una pequeña fracción glucídica (entre un 5 % y un 40 %) y una gran fracción proteica, que se unen mediante enlaces fuertes (covalentes).
Se diferencian de los proteoglucanos en que la parte glucídica no contiene ni ácido hialurónico ni sulfatos de condroitina.
Las principales son: las mucinas de secreción, como las salivales; las glucoproteínas de la sangre, como la protrombina y las inmunoglobulinas; las hormonas gonadotrópicas; algunas enzimas ribonucleasas y las denominadas glicoproteínas de las membranas celulares.
Los glucolípidos están constituidos por monosacáridos u oligosacáridos unidos a lípidos (ceramida). Generalmente se encuentran en la membrana celular. Los más conocidos son:
Texto:
Imagen:
Mira un ejemplo de lo que te pierdes
Categorias:
Etiquetas:
Deseja fazer um comentário? Registrar o Iniciar sessão
Si ya eres usuario, Inicia sesión
Adicionar ao Didactalia Arrastra el botón a la barra de marcadores del navegador y comparte tus contenidos preferidos. Más info...













Comente
0